 |
|||||||||||
 |
Stambrook's Lab |
||||||||||
|
|
|||||||||||
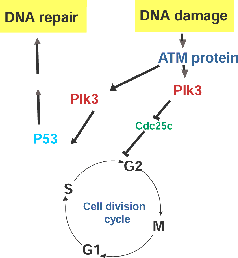
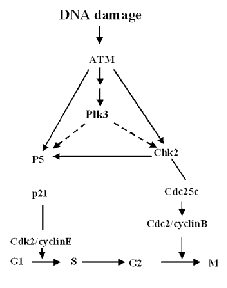
Peter J. Stambrook , Ph.D.: PLK3The polo kinases are a family of kinases that participate in cell cycle regulation, especially regulation of G2 and mitosis. There also is compelling evidence that some of the family members are involved in responses to stress such as damage to DNA and disruption of cytoskeleton. The polo kinase family was designated polo after the founding member, the Drosphila melanogaster polo gene and its protein product, which has a stretch of 28 amino acids, the polo box, that has been conserved from yeast to man. Disruption of polo in Drosophila embryos results in mitotic spindle defects, and micro- injection of antibody to human Plk1 (1 of the 3 polo kinases in mammals) into mamalian cells impair their ability to divide, resulting in a mitotic block, a reduction in centrosome size, and in immunoreactivity to MPM-2, a mitotic phospho-specific epitope. Mammals have at least three different, but related polo kinases (Plk1 through 3). We have recently shown that Plk3 is a participant in at least two cellular stress responses. Following either DNA damage or mitotic spindle disruption by nocodazole, Plk3 becomes phosphorylated and its kinase activity is enhanced. Its response to DNA damage is ATM dependent, whereas its response to spindle disruption is independent of ATM function. Plk3 appears to be centrally linked to multiple signaling pathways that respond to DNA damage. It co-immunoprecipitates with both P53 and with Chk2 as well as with members of the anaphase promoting complex (APC), a participant in a G2 ubiquitination pathway. It utilizes both P53 and ChK2 as kinase substrates, and it phosphorylates ChK2 at a site(s) different than ATM. Schematics of its potential role in DNA damage response and its potential interacting partners are presented in the figures. |
|||||||||||
 |
|||||||||||
 |
|||||||||||
|
As a first step to generate mice with single nucleotide polymorphisms (SNPs) in Plk3 that affect response to DNA damage and other environmental insults, it is necessary to recover the full length genomic DNA encoding Plk3. We have utilized the CITB mouse 129 BAC library from Research Genetics. The library covers the genome approximately 8 times, utilizing 221,000 independent clones with an average insert size of 130 kb. Each well of the super pool plate corresponds to clones from eight 384-well microtiter library plates. We have identified 5 Plk3 PCR positive clones from the super pool, using a primer set located within the first exon of Plk3. A second set of primers located 2 kb upstream verified the positive identification. Each positive pool from the super pool corresponds to a column of wells on one of the 6-plate pools. The positives from the plate pool PCR reaction determine which 384-colony spotted membrane to order from the vendor. The membranes obtained were hybridized using end-labeled oligonucleotides homologous to the beginning of the first exon. Seven positive clone addresses were identified and ordered, of these, five proved positive by verification with both sets of primers. The BAC clones are being expanded and the genomic Plk3 sequence is being subcloned. The nucleotide sequence and intron/exon organization has been deduced from the Celera database, and provides the necessary information for sub-cloning. Methods to introduce a single nucleotide change in the T loop and also in the ATP binding domain to eliminate kinase actively have been designed. |
|||||||||||
Interim ProgressThe DNAs of two independent strain 129 BAC clones were sequenced by the Kucherlapati Center, and yielded sequences that Plk3 genomic DNA that closely matched the Celera sequence. The Celera data base had indicated that the Plk3 gene was encompassed by a much larger gene. However, the sequencing data from our BAC clones did not bear this out. The restriction enzymes Nhe1 and Sal 1 which cut on the 5’and 3’of the gene respectively were selected for subcloning an 8.5kb fragment. The fragment was verified by Southern blot, but proved difficult to clone. We have tried to subclone this fragment into several recipient vectors, but to no avail. It appears to be very unstable for some unknown reason. We are now beginning to attempt to subclone it using a recombineering approach. The progress in this aspect of the Plk3 project has been disappointing, but we anticipate success using the recombineering approach. |
|||||||||||
|
|
|||||||||||
|
|
|||||||||||